In the past ten years, a series of skull fossils of Indarctos, Agriotherium and Ursavus have been discovered in the sediments of the Late Miocene Liushu Formation in the Linxia Basin, Gansu. A complete skull of Ursus has never been found and reported in the world before, so many questions about the origin, phylogeny and classification of Ursinae remain unanswered. In order to better understand the early members of bears, especially the nature of Ursus, academician Qiu Zhanxiang and others from the Institute of Vertebrate Paleontology and Paleoanthropology of the Chinese Academy of Sciences prioritized the skull and mandible fossils of Ursus discovered in the Linxia Basin for research. After in-depth research, the latest 2014 issue 3 of Vertebrata PalAsiatica published their research results as a cover article.

Figure 1. The fossilized skull of Ursula Dai and its head restoration, with the background being the late Miocene three-toed red soil of the Linxia Basin.
This ancestral bear fossil was discovered near Huaigou Village, Guanfang Township, Guanghe County, Linxia Hui Autonomous Prefecture. Fossils of the ancestors of Ursinae, especially those of the genus Ursus, are occasionally found in Miocene sediments in the Holarctic, but they are almost always represented by teeth and broken mandibles. The previous best specimen was the front half of a skull from the Greek island of Euboea, described in detail in 1947. Another well-preserved specimen is the skeleton of Ursavus orientalis from Shanwang, Linqu County, Shandong Province. It was reported by Academician Qiu Zhanxiang and others in 1985, but only some dental characteristics were described. Huaigou's skull is the first discovered and the only complete early Ursinidae fossil with a mandible.
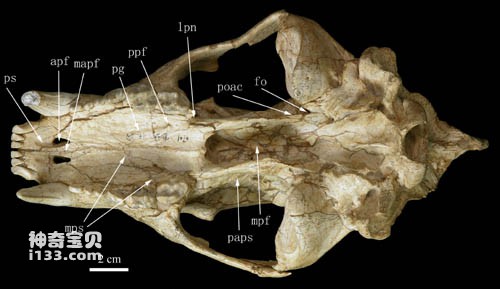
Figure 2 The ventral surface of the skull of Ancestral Dai's bear
In the same layer as the ancestral bear specimen at the Huaigou site, there are also fossils of the typical three-toed horse fauna in North China, including the original skunk, the hind cat, the saber-toothed tiger, the ferret hyena, the hyena, the three-toed horse, the great labial rhinoceros, The geological age of Chinese Elasmodon, Gulin and Proto-Chinese Antelope is approximately 8 million years ago.
Before the 1940s, Ancestral Ursus occupied a central position in the study of ancestral types of living bears. Since the late 19th century, nearly every bear-like carnivore researcher has believed that Ursus was the sole ancestor of living bears. The genus Ursavus was established by the German paleontologist Schlosser in 1899. He mainly based on Ursavus brevirhinus material from three locations in Austria and Poland (two incomplete mandibular ramus, one upper jaw with dentition, and some isolated teeth).
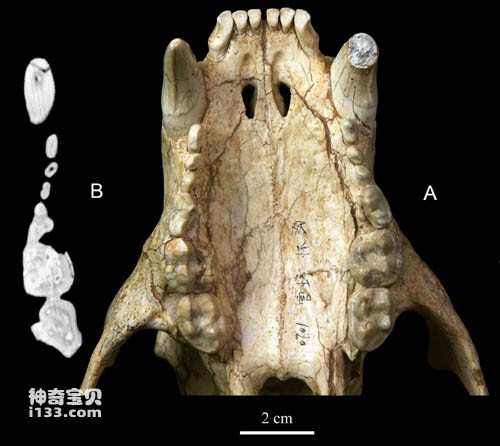
Figure 3 Upper cheek teeth of Ursus Dai's
Judging from Schlosser's description, the following characteristics of Ursus bear are of greater identification significance than those of living bears: small size, close to the size of wolves; canine teeth that are more laterally flattened, with obvious anterior and posterior ridges; premolars are always 4 pieces, with gaps between them. Except for the upper fourth premolar, the other premolars are simple and have no secondary cusps. The second and third premolars are double-rooted. The lower posterior tip of the upper fourth premolar is blade-shaped, its original tip is quite posterior, and the inner tooth belt is well developed. The coronal surfaces of the molars are strongly ruffled. The upper molars are rectangular, with length slightly greater than width. Each molar has two external cusps and two internal cusps. The medial base is strongly convex and the external tooth belt is well developed. The upper second molar has a short base. The lower first molar is large, and its lower triangular seat is much higher than the lower calcaneal seat. It has a strong but low inferior cusp and a small inferior inner cusp. The lower primary cusp of the lower second molar is low, but the lower posterior cusp is quite developed and there is no lower anterior cusp. The lower third molar is tumor-like.

Figure 4 The side view of the mandible of Ursus Dai's
Later, with the addition of new material, mainly isolated teeth, found in early and middle Miocene deposits, Ursus gradually became a rather mixed genus. So far, a total of 11 species have been described. Among paleontologists, the reasons for placing these species in the genus Ursus vary widely. At least two species, U. depereti and U. elmensis, were later formally excluded from the ancestral group. When Kretzoi established the new genus Agriarctos in 1942 based on some isolated teeth recovered from the latest Miocene strata in Hungary, he placed the holotype of U. depereti within its new genus and assigned it to the smaller species Agriarctos vighi. Mainly based on the identification characteristics of the lower first molar, such as the teeth being short and wide, with a strong forward-displaced lower posterior cusp and thus forming a rather closed lower triangular seat, the Suburban Panda was considered by Kretzoi to be between the Ancestral Bear and the Suburban Bear. -Transitional type between Indian bear lineages. In 1989, Qiu Zhanxiang and Qi Guoqin established a new genus and species Ailurarctos lufengensis belonging to the giant panda lineage based on the material originally attributed to the ancestral bear from the Lufeng hominin fossil site in Yunnan.
Ginsburg and Morales established a new genus Ballusia (Ballusia) based on the original Ursavus elmensis in 1998. Identifying features of this new genus include: generally smaller than Ursus ursus, a lingual crest formed by the protocusp and posterior cusp of the upper molar broadly separated from the labial anterior and posterior cusps, and a wider and more curved inner tooth band, The upper second molar is obviously oval in shape, with a short and wide base; the lower first molar is similar to that of Hemicyon and Plithocyon, both with a higher lower triangular base. They believed that Oriental Ancestral Bear was close to the root of this lineage, so Qiu Zhanxiang and Qiu Zhuding temporarily classified U. orientalis into the genus Ballusia in 2013.
Recently, Abella et al. created a new genus Kretzoiarctos in 2012. The material was a mandibular fragment with a left fourth premolar and a first molar found in Spain. It was originally described as a suburban panda. of a new species. Based on cladistic analysis, Abella et al. believe that Kratzoiarctos beatrix is the earliest known member of the giant panda lineage. Their subfamily Pandainae consists of two lineages: the Indian bear clade and the giant panda clade, with the genus Panda being initially separated from the other members of its clade. The cladistic analysis by Qiu Zhanxiang et al. showed that Panda clarkii is the sister group of two other genera, namely Suburban Panda and Eo-Panda. Since Eo-panda is definitely the earliest known ancestral type of the giant panda branch based on its tooth morphology, the Klebsi panda may not be the direct ancestor of the giant panda. In fact, the Klebsi panda does not possess the key traits of the giant panda: enlarged premolars, especially the front and rear accessory tooth tips and the short V-shaped lower triangular seat of the lower first molar. On the other hand, these characters as well as the small anterior accessory cusp and biprotocus of the upper fourth premolar can be found in Indian bears. Therefore, it is very likely that the Klebsi's panda is the ancestral type of the Indian bear lineage rather than the giant panda lineage.
Ginsburg and Morales concluded in 1998 that the earliest member of Ursinae was Ursus anluensis, which gave rise to the Ursus-Ursus lineage, while Ursus gave rise to Ursus and Indian bears, and Ursus and Ursus formed sisters. group relations. In this way, the original members of Ursinae include Ballusia (3 species), Ursavus (8 species), Agriarctos (3 species), Ailurarctos lufengensis and Kretzoiarctos beatrix, totaling 5 genera and 16 species.

Figure 5 Coronal surface of the mandible of Ursus Dai's
So far, no complete skulls and connected mandibles have been found in the early bears such as Anlu bears, Ursus ursus, Panda graminis, Suburban pandas and Eo-pandas. The best material is an Ancestor bear published in 1947. The front half of the skull and two fragments of the mandible published in 1887. Now this gap has been filled by fossil specimens from Linxia. Due to its latest existence (ca. 8 million years) and its highly advanced position among early bear ancestors, the Huaigou skull and mandible are very different from most species of the Early-Middle Miocene and Early Late Miocene. , because it has characteristics that are more advanced and close to those of living bears.
Comparative studies have shown that Huaigou's skull is different from all currently known bear ancestors, so it was designated as a new species: Ursavus tedfordi. The species name commemorates Richard Hall Tedford, an outstanding American Neogene biostratigrapher and paleontologist who passed away in 2011 and a well-known expert on carnivore fossils. Cephalogale minor is a large ancestral bear, intermediate in size, proportion and morphology between the extinct Cephalogale minor and the living black bear (Selenarctos thibetanus). Important features of Ursula daii include: its snout is short and wide compared to the back of its skull; its sagittal crest becomes very high posteriorly, extending behind the occipital surface; its cranium is quite narrow compared to its face; its hearing The vesicle is triangular in morphology with a short tubular external auditory canal; the mandible is robust; the premolars are strongly reduced to button-like shapes; the enamel folds on the cheek teeth are weak.
Comprehensive evidence shows that Ursus Dai's is undoubtedly the latest and most advanced species among the ancestors of living bears. However, because some of its characteristics of premature development are not seen in living bears, Ursula Dai's is likely to be a side branch, constituting the closest sister group to the living bear lineage.
animal tags:
We created this article in conjunction with AI technology, then made sure it was fact-checked and edited by a Animals Top editor.
you may also like

In a time long before the modern world knew the fearsome Komodo dragon, there existed another terrifying predator that roamed the ancient Earth: the Red Crocodile (Crocodilus ruber), a prehistoric beast with a striking resemblance to the Komodo dragon. These enormous reptiles were not only fearsome...

Scientists recently discovered the fossil of a 6.6-foot-tall (about 2-meter) penguin that lived in prehistoric Antarctica 34 million years ago, and its prototype was taller than most human males today. Paleontologists working at the Natural Sciences Museum in La Plata, Ar...

You’ve probably heard of the dodo bird, a now-extinct species that today can only be seen in museums. Popular culture often portrays it as a fat, clumsy bird, sometimes even in movies or cartoons. But beyond this caricature, how much do you actually know about the dodo—what it looked like, where i...

The discovery of ancient human fossils in Zhoukoudian made Bu Dasheng, who was deeply involved in the "Central Asian Fever", extremely excited. He decided to include the excavation and research of Zhoukoudian as a "related additional project" in his Central Asia expedition plan in cooperation...

When people think of bacteria, the first thing that people think of are probably those pathogenic bacteria that cause diseases and destroy lives. Therefore, it is inevitable to talk about bacteria. In fact, pathogenic bacteria are only a part of bacteria, and in the vast world of bacterial fa...

Modern argali sheep (Ovis) are widely distributed in a series of mountain ranges such as the Caucasus, Himalayas, Tibetan Plateau, Tianshan-Altai Mountains, Eastern Siberia, and the Rocky Mountains of North America. In Eurasia, argali fossils have only been found in small amount...

Regulations on the Protection of Paleontological FossilsChapter 1 General ProvisionsArticle 1 These Regulations are formulated in order to strengthen the protection of paleontological fossils and promote the scientific research and rational utilization of paleontological fossils.Article 2 Those enga...

At the end of the Devonian period, a certain kind of lobe-finned fish ventured out of the water and climbed onto land, becoming the earliest amphibians. Since then, vertebrates have entered a new world completely different from the environment they had lived in for hundreds of millions of yea...
Email: jsset668#gmail.com (change # to @) Please indicate your purpose of visit! Guangdong ICP No. 2022053326 XML| map| Chinese
